| 2.4. Genetische
Voraussetzungen für die Vererbung der Fellfarben Vererbung wird verstanden
als die Weitergabe genetischer Information. Ich möchte
(sehr grob) dazu einige Hinweise geben.
Die Mendelschen
Erbfaktoren:
In jahrelangen,
unzähligen Kreuzungsversuchen mit Erbsenpflanzen deckte
der Österreichische Mönch Gregor Mendel (1822-1884) die
Gesetze der Vererbung auf. Sie sind Grundlage der
heutigen Wissenschaft von den Genen. Mendel selbst wußte
damals noch nichts von Chromosomen und Genen. Dennoch ist
keines seiner Gesetze bisher widerlegt worden, sieht man
von einigen Ausnahmen ab, die z. b. auf der Möglichkeit
von Lethalfakktoren beruhen und die Ergebnisse verändern
können. Im Schatten der revolutionären Erkenntnisse
Darwins fanden seine bahnbrechenden Arbeiten damals kaum
Resonanz und wurden einfach vergessen. Erst 1900 wurden
sie von drei Forschern unabhängig voneinander
wiederentdeckt.
Auf Grund seiner
Kreuzungsversuche stellte Mendel drei Gesetze auf:
1. Gesetz der Dominanz
2. Gesetz von der Trennung der Merkmale
3. Gesetz der freien Kombination der Faktoren
(Unabhängigkeitsgesetz)
Zu 1.:
Es handelt sich hier um die
Kreuzung eines Merkmals (monohybride Kreuzung). Mendel
experimentierte mit Erbsensorten, die gelbe
und grüne Samen
hatten. Jede hatte sich selbst bestäubt und vererbte
rein weiter (P-Generation
= Elterngeneration). Bei der Kreuzung dieser Erbsen, die
er künstlich bestäubte, waren nur Erbsen mit gelben
Samen. Er führte diese Versuche zwei Jahre lang aus mit
immer gleichem Ergebnis. Er leitete daraus ab: Die F1-Generation
(oder Tochtergeneration) sieht
immer gleich aus, ist uniform. Er folgerte, daß das
Merkmal grün verdeckt sei und nannte dieses
unterdrückte Merkmal rezessiv
(oder zurückweichend). Den unterdrückenden Faktor für
gelb nannte er dominant (oder
vorherrschend).
Waren diese gelben Samen
der F1-Generation nun Mischlingssamen? Steckte das
Merkmal grün noch verdeckt darin? Es müßte dann bei
einer Kreuzung der Hybriden untereinander (bei
Selbstbestäubung) wieder sichtbar werden.
Die F2-Generation
(2. Tochtergeneration) ergab nun dreimal so viele gelbe
(dominante) wie grüne
(rezessive) Samen. Es ergab sich bei allen Versuchen
immer das Verhältnis 3:1.
Zu 2.:
Er stellte daraus das
zweite, das Spaltungsgesetz
auf und folgerte: wenn in der zweiten Tochtergeneration
die ursprünglichen Maerkmale der Elterngeneration wieder
auftauchen, so heiißt das, daß die Erbanlagen für
diese Merkmale in den Mischlingszellen nur vorübergehend
vereinigt sind und wieder getrennt werden können. In den
Keimzellen (Gameten)
kann nur der Faktor für das eine oder das andere Merkmal
enthalten sein
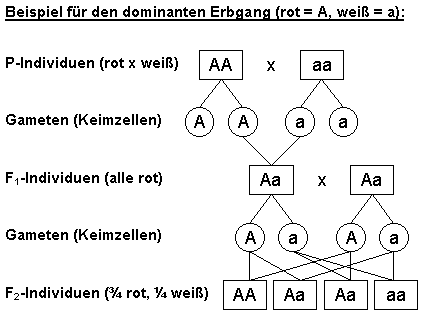
Ein
Drittel der dominanten rotblühenden Pflanzen (AA) und
die rezessiven weißblühenden(aa) vererben rein weiter,
sie sind homozygot,
während zwei Drittel der rotblühenden (Aa) gemischt
vererben, sie sind heterozygot. In
der F3-Generation spalten die Heterozygoten wieder gleich
auf: Ein Drittel der rotblühenden Pflanzen vererbt rein
weiter, zwei
Drittel spalten
wieder im Verhältnis 3:1..
P-Generation:
Die Erscheinungsfarbe (Phänotyp)
rot x weiß ist zu unterscheiden
vom
Genotyp AA x
aa, Gameten (A oder A) x (a oder a)
F1-Generation: Phänotyp 100%
rot, Genotyp 100% Aa
F2-Generation: Phänotyp 75% rot, 25% weiß, Genotyp 25% AA, 50% Aa, 25% aa
Zu
3.:
Damit ist der Erbgang eines
Merkmals untersucht. Je mehr Erbanlagen jedoch vorliegen,
desto komplizierter wird der Erbgang. Mendel führte ihn
mit zwei und mehr Merkmalspaaren durch. Er kreuzte
beispielsweise rein vererbende gelbe
runde (beide
dominant) und grüne
runzelige (beide
rezessiv) Samen (dihybride Kreuzung). In der
F1-Generation waren nur gelbe runde Samen.
In der F2-Generation erschienen aus 556 Pflanzen4
verschiedene Typen: 315 mit gelben
runden, 108 mit gelben
runzeligen, 101
mit grünen runden
und 35 mit grünen
runzeligen
Samen, d. h. rund im Verhältnis 9:3:3:1. Die Zahl der
Kombinationen ist 16.
Bei drei verschiedenen
Merkmalen ergeben sich acht verschiedene Typen, die Zahl
der Kombinationen erhöht sich auf 27. Bei 10
Merkmalspaaren ergeben sich 1024 verschiedene Pflanzen,
die 59 000 Kombinationsmöglichkeiten haben können.
(Alle diese Zahlen haben natürlich nur statistische
Bedeutung.)
Rück- und
Testkreuzung: Wenn bei der Kreuzung
zweier Pflanzen mit roten (AA) und weißen (aa) Blüten
3/4 der Blüten rot und 1/4 weiß sind, wäre zu prüfen,
welche dieser roten Blüten reinerbig (homozygot)
(AA) und welche gemischterbig (heterozygot)
(Aa) sind. Kreuzt man einen dominant mischerbigen Partner
(Aa) mit einem rezessiverbigen Partner (aa), so treten in
der der Nachkkommenschaft gemischterbige Rote (Aa) und
reinerbige weiße (aa) auf. Bei einem dominant
reinerbigen Partner wären alle mischfarbig rot (Aa).
Unvollständig
dominante und co-dominante Vererbung
Neben dem dominanten gibt es den intermediären
Erbgang, d.h. es können die
Merkmale der Eltern gleich stark einwirken. Aus einer
roten und einer weißen Blüte können beispielsweise in
der F1-Generation
rosa Blüten entstehen (unvollständig
dominant). Oder die Eigenschaften
beider Eltern können gemischt (co-dominant)
auftreten. Ein Beispiel wäre ein roan-farbenes Pferd aus
weißen und roten Eltern, bei dem rote und weiße Haare
nebeneinander gemischt auftreten .
Die Chromosomen:
Jede Zelle ist
durch eine äußere Membran in sich abgeschlossen.
Innerhalb dieser Zellwand befindet sich das zähflüssige
Zell- oder
Zytoplasma. Hier spielen sich die
Stoffwechselvorgänge der Zelle und die
Proteinbiosynthese ab, die vom Zellkern (Nukleus)
aus gesteuert werden. Eingebettet in den Zellkern sind
die Chromosonen. Sie
sind die Träger der genetischen Information.
Die Körperzellen besitzen
einen doppelten Chromosomensatz. Bei der Teilung einer
Körperzelle muß die Tochterzelle eine identischen
Chromosomenzahl erhalten. Die Chromosomen müssen sich
also verdoppeln. Diesen Prozeß bezeichnet man als Mitose-Zyklus
(mitos, gr. = Faden). Die Kernteilung vollzieht sich in
verschiedenen Phasen. Bei Beginn der Kernteilung bilden
sich die Chromosomen paarweise heraus, und zwar als lange
Fäden, die sich zunehmend verkürzen und bestimmte
einfache Formen annehmen. Sie bestehen der Form nach
jeweils aus identischen Doppelfäden (Chromatiden),
die sich umeinander drehen. An einer Einschnürungsstelle
werden sie zusammengehalten. Die Kernmembran löst sich
auf. Zwischen zwei Polen entwickeln sich Plasmafäden,
die den Chromosomen an den Einschnürungsstellen
anhaften. Gegen Ende der Phase teilen sich auch letztere.
Die Chromatiden rücken auseinander und wandern jeweils
zu einem der Pole, so daß die Chromosomensubstanz
gleichmäßig auf die beiden zukünftigen Kerne verteilt
ist. Es bilden sich zwei neue Zellen, die in der
Äquatorialebene voneinander getrennt werden.
Bei der Ei- oder Samenzelle
ist ein anderer Mechanismus festzustellen. Nur jeweils
ein Chromosom eines Paares gelangt in die Keimzelle (Gamete). Man nennt diewsen Vorgang Reifeteilung
oder Meiose,
die sich in zwei Schritten vollzieht. Im ersten werden
die Chromosomen getrennt und auf zwei Zellen verteilt,
Jede dieser Tochterzellen hat also einen einfachen
Chromosomensatz, der ein zufälliges Gemisch aus
mütterlichen und väterlichen Chromosomen ist. Die
zweite Reifeteilung gleicht einer Mitose, wobei ein
häufiger Stückaustausch stattfindet. Es entstehen vier
Keimzellen mit einem einfachen Chromosomensatz. Durch die
freie Kombination der Chromosomen und den
Stückeaustausch kommt es zu einer ungeheuer großen
Kombinationsmöglichkeit der Merkmale. Beim Mann gehen bei jeder Reifeteilung vier
Samen
hervor, bei der Frau bleibt nur
einer in der Eizellen, die anderen drei werden
abgeschnürt und sterben ab.
Bei
der Befruchtung verschmelzen die weibliche und die
männliche Keimzelle, so daß der ursprüngliche Satz
wieder erreicht wird. Bei der geschlechtlichen
Fortpflanzung sind die Keimzellen (Gameten)
des Vaters und der Mutter die
Träger des Erbmaterials. Die gesamte genetische
Information für ein neues Lebewesen ist erstmals in der
befruchteten Eizelle (Zygote) vorhanden.
Durch die Teilungen der lebendigen Zellen entsteht der
neue Organismus. Jede seiner Zellen enthält die gesamte
genetische Information.
Das Erbgefüge besteht aus
selbstständigen, voneinander trennbaren Einzelfaktoren,
die als Gengruppen auf den gleichen Chromosomen liegen.
Die Merkmale werden gekoppelt vererbt. Ausnahmen von
dieser Einschränkung werden durch den Stückeaustausch (Crossing-over)
ermöglicht, der mit statistischer Häufigkeit auftaucht.
Die Lebewesen haben
unterschiedlich viele Chromosomen. Die Zahl ist von ihrer
Organisationshöhe unabhängig. Der Mensch hat z.b. 46
(23 Paare), der Haushund 78
(39 Paare), der Einsiedlerkrebs 254 (127 Paare)
Chromosomen in den Körperzellen, jeweils paarweise
angeordnet. Davon stammt eines von der Mutter, eines vom
Vater.
Die Geschlechter
unterscheiden sich in einem Chromosomenpaar: das
weibliche, als X-Chromosom bezeichnet. und das männliche
Y-Chromosom. Es produziert in der Eizelle nur Keimzellen
vom X-Typus (XX), während die männlichen Keimzellen
sowohl X- als auch Y-Spermien produzieren (XY). Letztere
bestimmen also das Geschlecht des Nachkommen, je nachdem,
ob ein X- oder Y-Spermium auf das X-Chromosom der Eizelle
stößt.
Gene und Allele
Gene enthalten die
Information über ein Merkmal. Sie sind kleine Abschnitte
auf den Chromosomen und haben auf ihnen einen bestimmten
Platz, genannt locus.
Die beiden Partner eines Genpaares heißen Allele.
Wenn sie identisch sind, ist das Individuum homozygot
für ein Merkmal. Wenn zwei unterschiedliche Allele das
Paar bilden, ist das Individuum heterozygot: ein Allel
ist dominant, das andere rezessiv (Aa). Das dominante
Allel bestimmt das Aussehehn, das rezessive ist verdeckt,
jedoch vorhanden. Ein dominantes Gen wird mit einem
Großbuchstaben gekennzeichnet, ein rezessives mit einem
Kleinbuchstaben. Rezessive Gene können nur zkum Ausdruck
kommen, wenn sie homozygot (aa) auftreten.
Die DNS:
Die Chromosomen
bauen sich aus zwei Stoffgruppen auf: der DNS
(Desoxyribonucleinsäure), engl.
DNA, und den sie umgebenden Proteinen (Histonen).
Die haploiden
Keimzellen enthalten halb so viel DNS
wie die diploiden
Gewebezellen. Die Struktur kann man
sich vorstellen als eine eng gedrehte Doppelschraube, die
noch einmal um sich selbst gedrillt sein kann. Sie ist
durch angelagertes Histoneiweiß verdickt. Die DNS
hat die Fähigkeit, sich zu
verdoppeln. Dabei entsteht ein genaues Abbild (Replikation).
Es bilden sich zwei neue, identische Doppelstränge, die
zur einen Hälfte aus altem, zur anderen aus neuem
Material aus dem umgebenden Plasma des Zellkerns
bestehen. Die gleichbleibende Erhaltung des Erbgutes (und
die Weitergabe über die Keimzellen) ist somit durch die
Replikation gewährleistet. Die Selbstverdoppelung der
DNS geht der Längsspaltung der Chromosomen voraus. Die
Teile des Doppelstranges, die eine ein bestimmtes
Erbmerkmal betreffende Einheit darstellen, bezeichnet man
als Gene. Bei
homologen Chromosomen liegen sie jeweils an der gleichen
Stelle des Doppelstranges. Sie sind gleich, sofern sie
nicht durch Mutation verändert
wurden. Unter Gen versteht
man heute die Einheit der Funktion, d.h. die biochemische
Wirksamkeit, durch die die Herausbildung von Merkmalen
bewirkt wird.
Mutation:
Eine sprunghaft
auftretende Merkmalsänderung, die sich weitervererbt,
bezeichnet man als Mutation.
Es gibt drei Typen solcher Veränderungen: Genommutationen
(Erhöhung oder Verringerung der
Zahl der Chromosomen), Chromosomenmutationen
(Veränderungen der Struktur) und Genmutationen
(Veränderungen in sehr kleinen
Abschnitten, meist nur an einem Allel
eines Chromosomenpaares).
Mutationen lassen sich durch Chemikalien, am stärksten
aber durch UV-Bestrahlung stark steigern.
Allele Serien:
Während Mendel
noch von Faktoren sprach,
stehen heute dafür die Begriffe Allele
oder Gene.
Homologe Gene
liegen an der gleichen Stelle des Chromosoms (Allele).
Verändert eines durch Mutation seine Struktur und
Wirkungsweise, kann dieses mutierte Gen, bzw. neue Allel,
an die Stelle des ersten treten. Es kann eine ganze Serie
solcher Mutationen geben. Man spricht dann von multiplen
Allelen oder allelen
Serien. Jedes Allel liegt jeweils
an der gleichen Stelle (Locus),
die das ursprünglich unmutierte
Wildallel innehatte (mit
Pluszeichen, z.B. a+)
bezeichnet. Die mutierten Allele werden meist mit Zahlen
oder Buchstaben gekennzeichnet.
So wie sich die einzelnen
Faktoren gegenseitig unterdrücken, gilt dies auch
innerhalb der allelen Serien für die einzelnen Allele.
Im allgemeinen zeigt sich ein Farbverlust als Merkmal
jeweils unterlegen (rezessiv).
Die Allele verschiedener Serien können sich ebenfalls
gegenseitig unterdrücken, d.h. hypostatisch
sein, oder sie sind unterlegen, d.h.
epistatisch.
Neben einer Vielfalt
einfach dominanter oder rezessiver Faktoren vermuten die
Wissenschaftler heute bei der Farbvererbung der Hunde
mindestens vier solcher allelen Serien.
Quantitative
Vererbung:
Alle Faktoren
wirken in ihrer vielfältigen Kombination zusammen und
beeinflussen sich gegenseitig. Sie lassen nicht immer die
erwarteten Eigenschaften sichtbar werden, die das Erbbild
vorzugeben scheint. Bestimmte Faktoren werden nicht nur
durch ihre Gegenspieler, sondern auch durch die
Nachbarschaft anderer beteiligter Gene in ihrer
Entfaltung behindert oder unterstützt (Modifikatoren).
Die Intensität, mit der bestimmte Merkmale im Individuum
sichtbar werden, bezeichnet man als Expressivität
(Prägungsgrad)
der Gene.
Eigenschaften des Hundes,
wie Größe, Brustbreite, Länge, Schnelligkeit,
Fellansatz, Intelligenz etc. schwanken um einen
Mittelwert. An ihrer Ausprägung sind vermutlich viele
Gene auf verschiedenen Chromosomen beteiligt. Man nennt
diese Erscheinung ,quantitative
Merkmale‘. Ihre Weitergabe
kann vom Züchter nicht durch das Erkennen oder Verbinden
einzelner Gene (wie z.B. bei der Farbvererbung), sondern
nur durch die Zuchtwahl bestimmter Tiere gesteuert
werden, die die erwünschten Eigenschaften merklich
vorweisen. Deren Entwicklung ist jedoch zudem stark
abhängig von den Umweltbedingungen, d.h. von Klima,
Nahrung, Pflege, Zuneigung, Bewegung etc. Beim Welpen ist
daher noch kein endgültiges Urteil über die Erscheinung
des erwachsenen Hundes möglich.
|